
Genetic Code
mRNA template translation produces a protein product by converting genetic information into amino acid language. Among the amino acids that make up a protein, there are 20. Codons are triplets of nucleotides within the mRNA that represent one amino acid. The genetic code is a relationship between mRNA codons and their amino acid corresponding. A three-nucleotide code means that there are 64 combinations possible (43, each with four different nucleotides, within three different positions inside the codon).There are more codons for a given amino acid than there are amino acids, so the number is higher. Genetic degeneracy is the result of this redundancy. When choosing an amino acid to incorporate into a polypeptide, the codon's first two positions typically are crucial, but the codon's third position, also known as the wobble position, is less crucial. There is still a possibility of the amino acid being incorporated if the nucleotide in the third position is changed.
Nearly every individual has the same genetic code. A few species use different genetic codes for protein synthesis, but virtually all species do so, which strongly suggests that all life on the planet originated from a single source. Several unusual amino acids have been found in bacteria and archaea, including selenocysteine and pyrrolysine. Generally, UGA is a stop codon in the case of selenocysteine. The untranslated region of the mRNA can be used to encode selenocysteine through a stem-loop structure (known as a SECIS element or selenocysteine insertion sequence). A UAG codon stops translation at pyrrolysine. Pyrolysine is incorporated by a transfer RNA (tRNA) with the CUA anticodon.

Translation or protein synthesis
Translation involves several molecules and macromolecules in addition to the template mRNA. Ribosomes contain ribosomal RNA (rRNA) and polypeptides in different amounts depending on the organism. There are many similarities between the protein synthesis machinery of bacteria and human cells. The translation process involves mRNA, ribosomes, tRNAs, and other enzymes.Ribosomes
Ribosomes contain both catalytic ribozymes and structural RNAs, as well as large numbers of polypeptides. Approximately half of each ribosome is composed of mature rRNA. For example, the 70S ribosome is found in prokaryotes, and the 80S ribosome in eukaryotes, as well as the 70S ribosome in mitochondria and chloroplasts. A ribosome's dissociation from its small subunits during translation results in its reassembly when they are not synthesizing proteins. As a result, E. coli has the 70S (by no means additive) containing the small subunit (containing 16S rRNA) and the large subunit (containing 5S and 23S rRNA). In each ribosome there are two subunits: an 18S rRNA subunit in the small 40S subunit and a 5S, 5.8S, and 28S rRNA subunit in the large 60S subunit. In addition to binding the mRNA template, the small subunit attaches to tRNA.Several ribosomes are simultaneously translating each mRNA molecule, synthesising proteins from 5′ to 3′, and synthesizing polypeptides from the N terminus to the C terminus: reading the mRNA from 5′ to 3′. Polyribosomes, or polysomes, are structures containing multiple ribosomes accompanied by an mRNA. Bacteria and archaea synthesize numerous copies of polypeptides encoded by each transcribing transcript before transcriptional termination takes place since translation and transcription take place concurrently, forming polyribosomes.<
Both transcription and translation occur in the cytoplasm, since they occur in the same direction (5′-3′). Once the transcription is complete, the transcript is not processed. Once transcription is complete, transcription cannot be processed further. An environmental signal that triggers the production of new proteins can be responded to by prokaryotic cells very quickly. Eukaryotic cells cannot simultaneously perform transcription and translation. Eukaryotes produce polyribosomes as well, but these cannot form until the RNA synthesis process has been completed and the modified RNA has been transported out of the nucleus.<

Transfer RNAs
The transfer RNA (tRNA) is a structural RNA molecule found in the cytoplasm in many different species. The number of tRNA types varies from 60 to 90 in bacterial species. As each tRNA type binds to the codon coded in the mRNA template, the amino acid corresponding to the codon is added to the polypeptide chain. It is therefore tRNA molecules that translate RNA into protein. The tRNAs serve as translation adaptor molecules, which is surprising because they carry so much specificity in such a small package. The aminoacyl tRNA synthetase, ribosomal components, and mRNA interact with the tRNA molecule.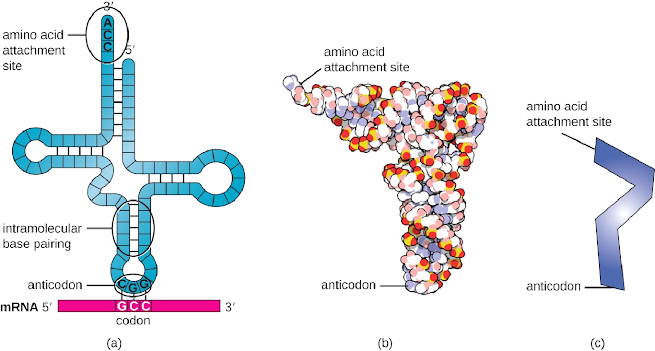
Mechanism
Initiation
The initiation of protein synthesis is marked by the formation of a protein synthesis initiation complex. A small 30S ribosome encodes the template mRNA, three initiation factors, and a special initiator tRNA carrying N-formyl-methionine (fMet-tRNA fMet). Methionine formylated on tRNA is present on the initiator tRNA that interacts with mRNA's AUG codon. A polypeptide chain synthesized by E. coli is inserted with fMet at its end (N terminus) because of its role in initiating polypeptide chains. The rRNA molecules that comprise ribosome interact with each other through complementary base pairing through the Shine-Dalgarno sequence downstream of the first AUG codon, or the ribosomal binding site AGG. During this interaction, the 30 S ribosomal subunit is anchored to the template mRNA at the correct location. The initiation complex is completed when the 50S ribosomal subunit binds to it.Similarly, the formation of initiation complexes differs between eukaryotes and prokaryotes:
- A different type of tRNA carries methionine, called Met-tRNAi, as the initiator tRNA.
- Eukaryotic initiation complexes are not localized to the Shine-Dalgarno sequence. Instead, they recognize the 5′ cap of eukaryotic mRNAs and proceed along their lengths after they reach the AUG start codon. A complex of Met-tRNAi, mRNA, and the 60S subunit is formed at this point.

Elongation
Neither prokaryotes nor eukaryotes exhibit elongation of translation in its basic form. As the 50S subunit binds with the intact ribosome, three functionally important ribosomal sites are formed. The A (aminoacyl) site is responsible for binding charged aminoacyl tRNA incoming into the ribosome. Proteinyl sites attach to amino acids that have formed peptide bonds with polypeptide chain formations but are still attached to their corresponding tRNAs. tRNAs that have been dissociated are released at the E site (exit), allowing them to be refueled by free amino acids. During initiation complex formation, tRNA molecules present in the initiation complex quickly enter the P site without first entering the A site. In this way, the tRNA matching the first codon after the AUG will be able to accept a free A site.During elongation, a ribosome moves a single codon at a time, a process known as translocation. Initially, charged tRNA enters at the A site, then shifts to the P site, and then leaves the E site once it has been taken out of the A site. Ribosomes undergo conformational changes in which they advance by three bases. The A-site tRNA and the P-site tRNA are attached by amino groups which form peptide bonds in the presence of peptidyl transferase, an RNA-based ribozyme located within the 50S subunit. Polypeptide chains are linked to amino acids bound to the P-site tRNA. A former P-site tRNA dissociates from an amino acid as the ribosome passes over the mRNA. Besides A site binding and translocation, GTP hydrolysis is also responsible for energy production. Specific elongation factors are responsible for GTP hydrolysis. Translation of a 200 amino acid protein takes just 10 seconds with the E. coli translation apparatus because each amino acid is added in 0.05 seconds.
Termination
If a nonsense codon (UAA, UAG, or UGA) occurs without a complementary tRNA, translation will be terminated. Prokaryotes and eukaryotes recognize these nonsense codons by their A site alignment, which causes amino acids at the P site to release from their tRNAs, releasing a polypeptide. The small and large ribosomal subunits separate from the messenger RNA and one another; they are subsequently recruited into another translation initiate complex.
Inhibitors
Translation (protein synthesis) is slowed or stopped by proteins synthesis inhibitors. Ribosomal enzymes are usually the targets of these inhibitors. Protein synthesis is carried out by both prokaryotes and eukaryotes using ribosomes. At the initiation, elongation, and termination of translation, these inhibitors work.Protein synthesis inhibitors are categorized according to their target organisms as follows:
- Acting on eukaryotes
- Acts on both prokaryotes and eukaryotes
- Acting on prokaryotes
Acting on eukaryotes
Anisomycin - The reaction of peptidyl transferase on ribosomes is blockedDiphtheria toxin - The enzyme inactivates eEF2, and eukaryotic elongation factor, by converting its diphthamide (a modified histidine) residue to ADP-ribose.
Cycloheximide - By inhibiting ribosome translocation (peptidyl transferase) in 80S eukaryotic ribosomes, Cycloheximide has anti-inflammatory properties.
Ricin - By depurination of a specific adenosine in 23S rRNA, this extremely toxic protein inactivates the 60S subunit of eukaryotic ribosomes.
Α-Amaniyin - Preferentially binds to RNA polymerase II to block mRNA synthesis.
Acts on both prokaryotes and eukaryotes
Actinomycin D - Actinomycin D binds to DNA, interferes with RNA polymerase movement, and inhibits RNA and protein synthesis.Puromycin - Puromycin is made by Streptomyces alboniger. An aminoacyl tRNA's 3' end has a very similar structure. The peptide bond formed between puromycin and ribosomal A produces peptididylpuromycin. It becomes dissociated from the ribosome shortly after its links to the carboxyl terminus of the peptide. Polypeptide synthesis is thus prematurely terminated. Not for clinical use since it is toxic to the host. Research is carried out with it, mainly cell culture research.
Acting on prokaryotes
Streptomycin - A soil actinomycete, Streptomyces griseus, is responsible for the production of streptomycin. The ribosome of bacteria is made up of the 30S, a small subunit of streptomycin. With relatively low concentrations, it can misread DNA and with higher concentrations, it can inhibit initiation.Erythromycin - A streptomycin-like antibiotic is manufactured by Saccharopolyspora erythraea (Streptomyces erythraeus). Erythromycin inhibits the synthesis of protein by binding to the 23S rRNA subunit of bacterial ribosomes. Binds to the ribosome exit channel, thereby inhibiting peptide chain elongation.
Tetracycline - Synthetic antibiotic derived from chlortetracycline via catalytic hydrogenation. The bacteria Streptomyces aurofaciens produces chlortetracycline. 30S ribosomal subunits bind to it. Ribosome site A is blocked from binding aminoacyl tRNA.
Chloramphenicol - Streptomyces Venezuela is the source of chloramphenicol. Antibiotic chloramphenicol binds reversibly to bacteria and is bacteriostatic. The binding of chloramphenicol to the aminoacyl tRNA binding site of the 50S subunit of the ribosome is meant to inhibit the activity of the peptidyl transferase.
Rifamycin - Protein synthesis is indirectly inhibited by rifamycin. RNA polymerase is inhibited by Rifamycin, which binds to it (Prevents RNA synthesis).
Get subject wise printable pdf notesView Here

No comments:
Post a Comment
Please don't spam. Comments having links would not be published.